丂丂堚揱巕偺懚嵼偩偗偑抦傜傟偰偄偰幚嵺偺惗棟婡擻偑晄柧側庴梕懱傪乽僆乕僼傽儞庴梕懱乿偲屇傃傑偡丅乽僆乕僼傽儞乿偲偼 乽屒帣乿偺偙偲偱丄恊乮忣曬偺壔妛揑幚懱乯偲偄偆傋偒婡擻惈偺乽惗懱撪儕僈儞僪乿乮儂儖儌儞側偳乯偑摨掕偝傟偰偄側偄偙偲偐傜丄偙偺傛偆偵屇偽傟傑 偡丅庴梕懱偺怴偟偄儕僈儞僪傪摨掕偡傞偨傔偵偼丄儕僈儞僪偺岓曗偲側傞朿戝側悢偺壔妛暔幙傗惗暔専懱傪曅偭抂偐傜妶惈應掕傪峴偄丄偟傜傒偮傇偟偵扵偟 偰偄偔偟偐偁傝傑偣傫丅偦偙偱嬌傔偰岠棪偺傛偄妶惈曽朄偑昁梫偲側傝傑偡丅摉尋媶幒偱偼庡偵乽庴梕懱亅G兛梈崌僞儞僷僋幙傪梡偄偨儕僈儞僪専嶕乿偲偄 偆曽朄傪梡偄偰偄傑偡丅
丂丂幚嵺偵儕僈儞僪扵偟傪偡傞庴梕懱偼偄傠偄傠偱偡丅
乮侾乯惗懱撪儕僈儞僪偑婛偵抦傜傟偰偄傞庴梕懱
- 婡擻偑暘偐偭偰偄傞庴梕懱偵寢崌偡傞暘巕傪扵偟弌偟偰丄怴偟偄栻偲偟偰巊偊側偄偐偳偆偐丄専摙偟傑偡丅丂
乮俀乯惗棟婡擻偑晄柧偺乽僆乕僼傽儞庴梕懱乿
- 婡擻偑暘偐傜側偄庴梕懱偼丄幚偼傑偩抦傜傟偰偄側偄怴偟偄儂儖儌儞傗恄宱揱払暔幙偵 懳偡傞庴梕懱偐傕偟傟傑偣傫丅惗懱撪偐傜偙傟傜偺僆乕僼傽儞庴梕懱傪妶惈壔偡傞暔幙傪扵偟弌偟偰丄怴偟偄惗懱撪挷愡婡峔偺夝柧偵敆傝傑 偡丅傑偨丄枴妎偺庴梕懱傗僼僃儘儌儞偺庴梕懱偲巚傢傟傞傕偺偵偮偄偰傕儕僈 儞僪偲側傞揤慠暔傪扵偟弌偟偰丄僸僩傗摦暔偺峴摦偑暘巕偵傛偭偰偳偺傛偆偵塭嬁傪庴偗傞偐傪尋媶偟偰傒偨偄偲巚偭偰偄傑偡丅
庴梕懱亅G兛梈崌僞儞僷僋幙傪梡偄偨儕僈儞僪専嶕丂
侾丏GPCR偲僴僀僗儖乕僾僢僩僗僋儕乕僯儞僌偺攚宨
G僞儞僷僋幙嫟栶庴梕懱乮GPCR乯偼嶰検懱G僞儞僷僋幙偲嫟栶偟偰摥偒丄尰嵼巊梡偝傟偰偄傞椪彴栻偺栺俆妱埲忋偑GPCR傪僞乕僎僢僩偵偟偰偄傞 偲偄傢傟偰偄傞丅GPCR偺儕僈儞僪専嶕偼岞揑尋媶婡娭偩偗偱側偔憂栻傪傔偞偡惢栻夛幮傗儀儞僠儍乕婇嬈側偳偑惙傫偵峴偭偰偄傞偑丄偦偺妶惈應掕朄偺 傎偲傫偳偼摦暔攟梴嵶朎偵懳徾偲偡傞GPCR傪敪尰偝偣丄儕僈儞僪岓曗偲側傞暔幙傪嶌梡偝偣偨帪偵婲偙傞嵶朎偺曄壔傪娤應偡傞偙偲傪婎杮偵偟偰偄傞丅 懡偔偺曽朄偑奐敪偝傟偨拞偱傕偭偲傕惉壥傪偁偘丄峀偔梡偄傜傟偰偄傞曽朄偼傾僑僯僗僩仺GPCR仺G僞儞僷僋幙乮摿偵Gq宯乯仺PLC仺IP3仺嵶朎 撪僇儖僔僂儉擹搙忋徃偲偄偆堦楢偺曄壔偱婲偙傞曄壔傪丄嵶朎偵摫擖偟偨Fura-2傗Fluo-3側偳偺寀岝偺曄壔偱娤應偡傞曽朄偱偁傞丅崱擔偱偼 Gi側偳偵嫟栶偟偨GPCR偵懳偡傞姶搙傪忋偘傞偨傔丄G16僞儞僷僋幙傗Gq/i僉儊儔僞儞僷僋幙傪GPCR偲嫟敪尰偝偣偨嵶朎傪梡偄傞偙偲偑懡偄 偑丄偙偺曽朄偺栤戣揰傕巜揈偝傟偰偄傞丅傑偨庴梕懱敪尰嵶朎偵傕偲傕偲懚嵼偡傞撪嵼惈庴梕懱偺岠壥偑丄妶惈應掕偺寢壥傪崿棎偝偣傞傕偲偲側傞丅
変乆偼婛懚偺僗僋儕乕僯儞僌宯偺栤戣揰傪崕暈偡傞傋偔丄廬棃偲偼堎側傞曽朄偱GPCR儕僈儞僪偺僴僀僗儖乕僾僢僩僗僋儕乕僯儞僌偑壜擻偵側傞曽朄傪 柾嶕偟偨丅摿偵Gi/o偼擼偱偺敪尰偑嫮偄偵傕偐偐傢傜偢偦傟偲嫟栶偟偨GPCR偺妶惈應掕偼忋婰偺曽朄傪娷傔婛懚偺曽朄偑斾妑揑嬯庤偲偡傞偺偱丄 Gi/o偵嫟栶偟偨GPCR偵廳揰傪偍偄偨儕僈儞僪専嶕宯偺峔抸傪栚巜偟偨丅
丂
俀丏庴梕懱亅G兛梈崌僞儞僷僋幙偵傛傞儕僈儞僪専嶕偺幚嵺
丂偝傑偞傑側専摙偺寢壥丄変乆偼庴梕懱偲G僞儞僷僋幙兛僒僽儐僯僢僩偲偺梈崌僞儞僷僋幙傪崺拵嵶朎(Sf9)偱挷惢偟丄偦偺枌夋暘傪梡偄偰傾僑僯僗 僩埶懚惈[35S]-GTP兞S寢崌妶惈傪應掕偡傞宯傪棙梡偡傞偙偲偲偟偨.梈崌僞儞僷僋幙傪娷傓枌夋暘偵傾僑僯僗僩傪壛偊傞 偲庴梕懱偵偮偯偄偰G僞儞僷僋幙兛僒僽儐僯僢僩晹暘偑妶惈壔偝傟傞偑丄G兛偵偼GTPase妶惈偑偁傞偺偱妶惈壔偵嵺偟偰G兛僒僽儐僯僢僩偐傜偺 GDP偺夝棧偑懀恑偝傟GTP偑寢崌偡傞丅変乆偺宯偱偼GTP偺偐傢傝偵[35S]-GTP兞S傪梡偄丄偙偺庢傝崬傑傟偨[35S]-GTP兞S 偺検傪僔儞僠儗乕僔儑儞僇僂儞僞乕偱應掕偟丄掕検壔偡傞丅
丂Sf9嵶朎傊偺梈崌堚揱巕偺摫擖偵偼慻姺偊懱僶僉儏儘僂僀儖僗傪梡偄傞偑堦楢偺夁掱偼僉僢僩壔偝傟偰偄偰娙曋偱偁傞丅儕僈儞僪僗僋儕乕僯儞僌偼 96寠僾儗乕僩傪梡偄偰峴偆偺偱丄庤嵺傛偔弨旛偟偰偍偗偽慡峴掱傪侾恖偱幚尡偟偰傕俀侽枃偺僾儗乕僩傪張棟偡傞偺偼乮應掕傪彍偒乯俀帪娫埲撪偵廔椆偱 偒傞丅
丂
俁丏僗僋儕乕僯儞僌宯偺昡壙
丂変乆偺曽朄偺挿強偼丄傑偢扨弮側binding assay傪偡傞偩偗偱偁傝斀墳傗嶌嬈偑娙扨側偙偲偱偁傞丅傑偨偙偺曽朄偱偼懠偺曽朄偲堎側傝丄戝検偺攟梴嵶朎傪宲戙偟懕偗傞昁梫偑側偄丅Sf9偼悢 儕僢僩儖偺僗働乕儖偱晜梀攟梴壜擻偱丄挷惢偟偨枌夋暘傪搥寢偝偣偰偍偗偽偄偮偱傕慡偔摨偠斾妶惈偺枌夋暘傪偡偖偵巊梡偱偒傞丅堦搙侾亅俁儕僢僩儖掱搙 偺攟梴傪峴偊偽丄偦偺枌夋暘偺検偱枅擔僩僢僾僇僂儞僩偺張棟検偄偭傁偄傑偱傾僢僙僀偟偨偲偟偰傕丄敿擭埲忋偼廫暘偵應掕偟懕偗傞偙偲偑偱偒傞丅攟梴嵶 朎傪巊梡偡傞応崌偵嵶朎偺宲戙傗庴梕懱偺埨掕揑側敪尰偵婥傪巊偭偨傝僗僋儕乕僯儞僌偺僗働僕儏乕儖偵崌傢偣偰嵶朎傪憹傗偟偨傝偡傞斚嶨偝傪峫偊傞偲丄 偙偺棙揰偼尰幚揑偵偼偐側傝堄枴偑戝偒偄偟埨怱偱偁傞丅僼儖傾僑僯僗僩偲僷乕僔儍儖傾僑僯僗僩傪嬫暿偟偨儕僈儞僪偺夝愅偑壜擻偱偁傞偙偲偼栻棟妛揑偵 偼廳梫偱偁傞丅Sf9嵶朎偺撪嵼惈庴梕懱偵傛傞妶惈應掕忋偺暃嶌梡偼偙傟傑偱曬崘偝傟偰偄側偄丅
丂堦曽抁強偲偟偰偼Gq偲嫟栶偡傞庴梕懱偵偮偄偰偆傑偔偄偐側偄偙偲偱偁傞丅偟偐偟歁擕椶攟梴嵶朎偱庴梕懱亅Gq兛梈崌僞儞僷僋幙傪敪尰偝偣偨応崌 偵偼[35S]-GTP兞S 偺庢崬傒検偺忋徃偑尠挊偵曬崘偝傟偰偄傞偺偱丄崱屻Sf9偱傕忦審傪専摙偡傟偽Gq傊偺揔梡偑壜擻 偵側傞偲峫偊偰偄傞丅傑偨嵶朎傪巊偆応崌偺傛偆側庴梕懱埲壓偺僔僌僫儖揱払僇僗働乕僪偵傛傞怣崋偺憹暆偑婜懸偱偒側偄丅偟偨偑偭偰傾僑僯僗僩偺専弌壓 尷擹搙偼儕僈儞僪偲庴梕懱偺娫偺Kd抣偍傛傃[35S]-GTP兞S偺斾妶惈偱婯掕偝傟傞丅偙傟偼応崌偵傛偭偰偼婛懚偺曽朄偺 10攞掱搙姶搙偑埆偄偲昡壙偡傞偙偲傕偱偒傞偑丄攟梴嵶朎偺応崌偼撪嵼惈庴梕懱偺岠壥傪妋擣偡傞偨傔偵帋椏傪旓傗偡偙偲側偳傪崌傢偣偰峫偊傞傋偒偱偁 傞丅
丂
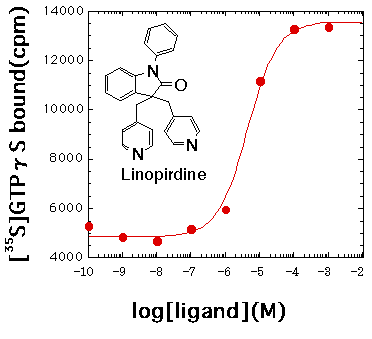
丂変乆偼婛偵偙偺曽朄傪梡偄偰僸僩僎僲儉忣曬偐傜怴婯偵摨掕偟偨hGPCR48偑5-oxo-ETE庴梕懱偱偁傞偙偲丄Linopiridine偑 hGPCR19偺傛偄傾僑僯僗僩偱偁傞偙偲側偳傪敪尒偟偨丅
丂
仾丟hGPCR19偼Linopiridine偵傛傝妶惈壔偝傟傞偙偲傪hGPCR19-Gs兛梈崌僞儞僷僋幙傪梡偄偰帵偟偨寢壥
丂